支气管哮喘发病机制的新认识
2018/12/24
费凡 吉宁飞 黄茂
江苏省人民医院(南京医科大学第一附属医院) 210029
江苏省人民医院(南京医科大学第一附属医院) 210029
哮喘是一种复杂的、异质性的慢性气道炎症性疾病,其发病机制尚未完全明确。最早在1878年发现了肥大细胞和嗜酸细胞,1986年定义了小鼠中的Th1和Th2细胞,直到1999年提出了哮喘表型,2003年鉴定了调节性T细胞(Treg)、Th17、Th9和固有淋巴细胞(ILC)的作用机制。近年来,随着对哮喘的不断深入研究,其发病机制有了更多的新进展。
1 哮喘的遗传学进展
1.1 哮喘的表型、内型与基因型 越来越多的研究证实,哮喘的表型、内型与基因型密切相关。有学者[1]提出哮喘的表型与内型之间的联系,2型免疫应答优势型哮喘可分为3个主要的表型:嗜酸细胞性炎症、变应原致敏(存在抗原特异性IgE)、气道高反应性及重塑。在儿童哮喘的不同表型中,同样存在不同的内型(endotype)[2]。在嗜酸性哮喘儿童中,通常观察到增厚的基底膜,上皮细胞死亡和微血管形成。而在嗜中性粒细胞哮喘儿童的痰液细胞内表现出基因粘液病毒抗性1,寡聚腺苷酸合成酶和维甲酸的过度表达[3]。此外,有学者[4]研究了哮喘和COPD有关的内型和相应的基因型,并提出,一些临床表型可以在一个患者中重叠,相同的临床表型可能来自不同的内型和基因型。因此,未来的治疗方案可能针对各种由不同的病理生理机制定义的内型,如2型炎症,病毒感染的易感性的增加,肺发育损伤,细菌克隆(表1)。

1.2 哮喘的遗传学进展
1.2.1 遗传学证据:哮喘的发生具有家族聚集性特点,同卵双生的双胞胎共同发生哮喘的比率高于异卵双生的双胞胎,哮喘的遗传度约为60%[5]。哮喘是多基因遗传病,目前已陆续发现了数百个哮喘易感基因,这些基因与特应性(atopy)、气道高反应性(AHR)、炎症介质(如细胞因子、趋化因子、生长因子等)的产生、Th细胞亚群比例等均有相关。
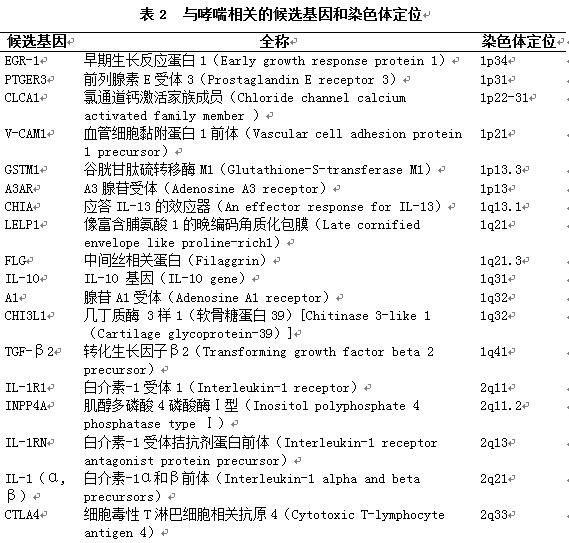
1.2.2 常用研究策略及候选基因:主要包括全基因组关联研究、候选基因策略和定位克隆策略。目前已知的哮喘的遗传性进展主要来自全基因扫描及候选基因(Candidate genes)的研究结果。近年来研究的相关热门候选基因见表2[6]。基于GWAS研究[7],发现有意义的候选基因见表3。







1.2.3 基因多态性 除了候选基因外,候选基因的基因多态性(SNPs)与哮喘密切相关。近年来相关的基因多态性研究热点有以下内容:细胞因子及其受体,趋化因子及其受体,β2肾上腺素受体,血管紧张素I转化酶,解聚素和金属蛋白酶域33,血清类黏蛋白1样蛋白3,谷胱甘肽硫转移酶。在过敏性哮喘中,目前较为认可的基因多态性包括:HLA DQ、HLA DPB1、IL4、IL13、TEBT、IL10&TGF-Bata、ACE等[7]。还有更多的基因多态性等待研究。
2 哮喘的表观遗传学进展
遗传和环境因素的组合被认为影响炎症消退或进展的重要因素,因此表观遗传修饰在环境和遗传因素之间的调控尤为重要。哮喘发生的表观遗传学调控包括DNA甲基化、组蛋白修饰、非编码RNA调控等[8]。
2.1 DNA甲基化 改变DNA甲基化状态可能导致细胞因子及转录因子的基因表达差异。在Dp/Df激活的哮喘患者中,未甲基化的IL-4和I;-4的表达水平增高相关[9]。Somineni等人的研究发现在空气污染地区出生的儿童与健康的兄弟姐妹和对照者相比较,TET1启动子的CpG位点显示出低甲基化[10]。Murphy等人的单卵双生子的哮喘研究显示了不同的甲基化模式与儿童哮喘有关,HLX基因下游的CpGs不同的甲基化模式提示了哮喘可能的发病机制[11]。在一项哮喘患者的全基因组分析研究中[12]发现,气道中的DNA甲基化在介导遗传变异对哮喘风险和临床过程的影响方面起核心作用。
2.2 组蛋白修饰 组蛋白修饰可以对基因表达具有活化和抑制功能。每个组蛋白残基的修饰程度和修饰的组合和位置对基因表达和组蛋白结构的影响可能有显着差异[13]。调控组织蛋白PTM的许多酶在真核生物中大部分保守。其表达受到遗传和环境因素的影响。例如,严重哮喘儿童的被动吸入烟雾大大降低了组织蛋白脱乙酰酶(HDAC)2在肺泡巨噬细胞中的表达。HDAC2可以在组蛋白3和组蛋白4上赖氨酸残基脱乙酰化[14]。 HDAC2由皮质类固醇上调,从而抑制炎症基因表达和减轻肺损伤。组蛋白修饰基因的治疗潜力具有很强的证据基础,特别是HDAC2在哮喘中的作用。
2.3 非编码RNA miRNA是一类长19~25个核苷酸的小分子非编码RNA,它不仅参与了DCs的表型分化,而且还参与了幼稚T淋巴细胞的分化,参与哮喘气道炎症和气道重塑的调控,microRNAs被证明是未来哮喘治疗的目标[15]。Levanen等[16]对哮喘患者和健康者的支气管肺泡灌洗液中miRNA进行差异性芯片检测,发现有24个miRNA表达存在明显差异,其中主要包括let-7家族成员。let-7 miRNA可抑制IL-13表达,降低Th2炎症[17]。
3 哮喘的气道炎症、气道重塑和气道高反应性
3.1 哮喘的气道炎症 不同的哮喘内型具有不同程度的炎症、气道反应性、黏液分泌及气道重塑,可通过上皮细胞及免疫细胞等介导[18]。在过敏或IgE介导的哮喘的典型模型中,各种环境暴露触发气道上皮细胞和引发免疫应答的树突状细胞相互作用。气道上皮通过模式识别受体识别蛋白酶活性变应原或微生物,导致多种细胞因子、核苷酸和脂肪酸代谢物的表达,随后发送信号给负责招募的TH1或TH2细胞因子,嗜中性粒细胞和嗜酸性粒细胞的免疫细胞[19]。树突细胞处理来自这些环境暴露的抗原,并将其呈递给T细胞,引发TH2型淋巴细胞的克隆扩增。TH2型细胞的克隆扩增导致IL-4,IL-13和IL-5的形成以及B细胞的分类转换,产生IgE。此外,TH2细胞因子上调肥大细胞和嗜酸性粒细胞增殖,活化的肥大细胞和嗜酸性粒细胞产生额外的TH2细胞因子,使气道炎症持续。来自哮喘的支气管活检显示,哮喘患者存在嗜酸性粒细胞,活化的肥大细胞和Th2细胞为主的T细胞浸润。由于增生和肥大,导致气道上皮发生结构性改变,包括上皮下的胶原沉积和气道平滑肌增厚,新生血管数量以及粘液增生也有增加[20]。
3.2 气道重塑 气道重塑的特征是存在不同的结构改变,例如粘液的化生,网状层状增厚,血管生成增加,上皮下纤维化和平滑肌肥大、增生。这些结构修饰都是由不同的免疫学相关的介质或细胞机制。与气道重塑相关的细胞主要包括T细胞,嗜酸性粒细胞,肥大细胞,成纤维细胞和其他上皮细胞。近年来研究显示,气道上皮细胞在气道重塑起到重要作用[21]。上皮间充质营养单元(EMTU)是上皮细胞和间充质细胞之间的成纤维细胞层。这些成纤维细胞能够分化为肌成纤维细胞,作为对一些局部刺激的反应,并分泌不同的细胞介质和蛋白质,影响上皮细胞,平滑肌细胞,血管和神经末梢的动态结构。支气管上皮间质转化 (epithelial-mesenchymal transformation, EMT)通过上皮-间充质营养单位的可变活化,可能导致不同哮喘表型的出现,从而提供更有针对性的治疗方法[22]。
3.3 气道高反应性 气道高反应性(AHR)表现为气道对各种刺激因子出现过强或过早的收缩反应,是哮喘发生发展的重要因素。Nassenstein[23]研究提示,神经生长因子可能是调节哮喘神经-内分泌-免疫网络失衡机制中的启动因素,它由炎症部位的细胞产生,引起平滑肌收缩,调节白细胞产生促炎分子,促使气道感觉神经增生、敏感化,导致气道高反应性。近年来,发现多种产物可能和气道高反应性有关,如p22phox[24],白细胞介素33[25],α5β1整联蛋白[26]等。
4 哮喘的其他相关机制
4.1 职业性哮喘的发病机制 职业性哮喘(OA)是所有工业化国家最常见的工作型肺病之一。已经发现许多因素有助于化学诱导的OA的发病机制,包括先天免疫机制,以及上皮损伤,气道重塑,氧化应激,神经源性炎症和遗传风险因素的非免疫机制。与OA易感性增加相关的基因包括HLA II类基因和与先天免疫,Th2免疫和抗氧化酶基因相关的基因[27]。
4.2 儿童肥胖与哮喘的可能机制 AHR是哮喘的主要特征。当肥胖个体的肺容积减少时,气道直径相应减少,随着时间的推移,它可以干扰平滑肌功能并增加气道阻塞和高反应性。有针对儿童和成人哮喘的前瞻性研究报告[28]显示,肥胖成年人的肺功能明显下降,儿童则无此结果。可见肥胖儿童和哮喘的关系尚未明确,表明在儿童肥胖研究中需要更多的调查评估。
4.3 吸烟与哮喘的相关机制 哮喘中的吸烟常常存在,并且常常伴随着症状恶化,肺功能下降加快,入院次数增加,哮喘严重程度更高,哮喘控制较差,对皮质类固醇反应降低。此外,它与呼吸道壁粘膜下层中嗜酸性粒细胞数量减少和肥大细胞数量增加有关。吸烟哮喘患者的上皮厚度和杯状细胞增生增加证明气道重塑增加。烟草诱发气道炎症改变和哮喘重塑的发病机制是复杂的,很大程度上是未知的。减少皮质类固醇反应的潜在机制也是未知的[29]。
4.4 哮喘急性发作的可能机制 在急性哮喘中使用诱导痰的气道炎症研究表明,哮喘急性发作具有嗜中性粒细胞和嗜酸性粒细胞混合物的异质炎症浸润,这种炎症浸润的模式与过敏原诱发的哮喘模型不同。这表明急性哮喘的发病机制与慢性疾病的发病机制不同[30]。T细胞活化是急性重症哮喘的重要特征,表现为外周血中T细胞标志物增加,以及致死性哮喘病例组织中活化(CD25 +)CD8细胞数量增加。氧化应激是急性重症哮喘的另一个关键,急性哮喘中标记的粒细胞流入和激活伴随着氧自由基生成增加,宿主抗氧化防御被破坏,并导致脂质和蛋白质的氧化[51]。
5 总结与展望
哮喘是多基因病,多基因病的复杂性导致哮喘的遗传学进展较慢,目前仅有相关的候选基因。表观遗传学的出现,使我们能深入了解哮喘发病的基因调控机制。哮喘的发病机制复杂,气道炎症、重塑和AHR等只是其理论的一部分,相关的炎性细胞、炎性介质等形成了复杂的网络,如何全局把握其发病机制,成为研究的难点。相信随着生物学、免疫学、遗传学以及表观遗传学等的进展,在不久的将来,将会有更大的研究突破。
参考文献:
[1] MURARO A, LEMANSKE JR R F, HELLINGS P W, et al. Precision medicine in patients with allergic diseases: Airway diseases and atopic dermatitis—PRACTALL document of the European Academy of Allergy and Clinical Immunology and the American Academy of Allergy, Asthma & Immunology [J]. Journal of Allergy and Clinical Immunology, 2016, 137(5): 1347-58.DOI:10.1016/j.jaci.2016.03.010
[2] CHOI H, SONG W-M, ZHANG B. Linking childhood allergic asthma phenotypes with endotype through integrated systems biology: current evidence and research needs [M]. Reviews on Environmental Health. 2017: 55. DOI:10.1515/reveh-2016-0054
[3] DA SILVA J, HILZENDEGER C, MOERMANS C, et al. Raised interferon-β, type 3 interferon and interferon-stimulated genes – evidence of innate immune activation in neutrophilic asthma [J]. Clinical & Experimental Allergy, 2017, 47(3): 313-23.DOI:10.1111/cea.12809
[4] HIZAWA N. Clinical approaches towards asthma and chronic obstructive pulmonary disease based on the heterogeneity of disease pathogenesis [J]. Clinical & Experimental Allergy, 2016, 46(5): 678-87.DOI:10.1111/cea.12731
[5] THOMSEN S F, VAN DER SLUIS S, KYVIK K O, et al. Estimates of asthma heritability in a large twin sample [J]. Clinical & Experimental Allergy, 2010, 40(7): 1054-61.DOI:10.1111/j.1365-2222.2010.03525.x
[6] BIJANZADEH M, MAHESH P A, RAMACHANDRA N B. An understanding of the genetic basis of asthma [J]. The Indian Journal of Medical Research, 2011, 134(2): 149-61.
[7] LEE J-U, KIM J D, PARK C-S. Gene-Environment Interactions in Asthma: Genetic and Epigenetic Effects [J]. Yonsei Medical Journal, 2015, 56(4): 877-86.DOI:10.3349/ymj.2015.56.4.877
[8] ALIZADEH Z, MORTAZ E, ADCOCK I, et al. Role of Epigenetics in the Pathogenesis of Asthma [J]. 2017, 2017, 10.DOI:
[9] KWON N-H, KIM J-S, LEE J-Y, et al. DNA Methylation and the Expression of IL-4 and IFN-γ Promoter Genes in Patients with Bronchial Asthma [J]. Journal of Clinical Immunology, 2008, 28(2): 139-46.DOI:10.1007/s10875-007-9148-1
[10] SOMINENI H K, ZHANG X, BIAGINI MYERS J M, et al. Ten-eleven translocation 1 (TET1) methylation is associated with childhood asthma and traffic-related air pollution [J]. Journal of Allergy and Clinical Immunology, 2016, 137(3): 797-805.e5.DOI:10.1016/j.jaci.2015.10.021
[11] MURPHY T M, WONG C C Y, ARSENEAULT L, et al. Methylomic markers of persistent childhood asthma: a longitudinal study of asthma-discordant monozygotic twins [J]. Clinical Epigenetics, 2015, 7(130.DOI:10.1186/s13148-015-0163-4
[12] NICODEMUS-JOHNSON J, MYERS R A, SAKABE N J, et al. DNA methylation in lung cells is associated with asthma endotypes and genetic risk [J]. JCI Insight, 2016, 1(20).DOI:10.1172/jci.insight.90151
[13] KIDD C D A, THOMPSON P J, BARRETT L, et al. Histone Modifications and Asthma. The Interface of the Epigenetic and Genetic Landscapes [J]. American Journal of Respiratory Cell and Molecular Biology, 2015, 54(1): 3-12.DOI:10.1165/rcmb.2015-0050TR
[14] KOBAYASHI Y, BOSSLEY C, GUPTA A, et al. Passive Smoking Impairs Histone Deacetylase-2 in Children With Severe Asthma [J]. Chest, 2014, 145(2): 305-12.DOI:10.1378/chest.13-0835
[15] KAI W, QIAN X U, QUN W U Z. MicroRNAs and Asthma Regulation [J]. Iranian Journal of Allergy, Asthma and Immunology, 2015, 6.DOI:
[16] KUMAR M, AHMAD T, SHARMA A, et al. Let-7 microRNA–mediated regulation of IL-13 and allergic airway inflammation [J]. Journal of Allergy and Clinical Immunology, 2011, 128(5): 1077-85.e10.DOI:10.1016/j.jaci.2011.04.034
[17] GREENE C M, GAUGHAN K P. microRNAs in asthma: potential therapeutic targets [J]. Current Opinion in Pulmonary Medicine, 2013, 19(1): 66-72.DOI:10.1097/MCP.0b013e32835a5bc8
[18] OLIN J T, WECHSLER M E. Asthma: pathogenesis and novel drugs for treatment [J]. BMJ : British Medical Journal, 2014, 349. DOI: 10.7196/SAMnew.8207
[19] HARPER R W, ZEKI A A. Immunobiology of the Critical Asthma Syndrome [J]. Clinical Reviews in Allergy & Immunology, 2015, 48(1): 54-65.DOI:10.1007/s12016-013-8407-6
[20] ISHMAEL F T. The Inflammatory Response in the Pathogenesis of Asthma [J]. The Journal of the American Osteopathic Association, 2011, 111(11_suppl_7): S11-S7.DOI:
[21] MANSO L, RECHE M, PADIAL M A, et al. Diagnostic tools assessing airway remodelling in asthma [J]. Allergologia et Immunopathologia, 2012, 40(2): 108-16.DOI:10.1016/j.aller.2011.11.002
[22] BITTAR H E T, YOUSEM S A, WENZEL S E. Pathobiology of Severe Asthma [J]. Annual Review of Pathology: Mechanisms of Disease, 2015, 10(1): 511-45.DOI:10.1146/annurev-pathol-012414-040343
[23] NASSENSTEIN C, KUTSCHKER J, TUMES D, et al. Neuro–immune interaction in allergic asthma: role of neurotrophins [J]. Biochemical Society Transactions, 2006, 34(4): 591.DOI:10.1042/BST0340591
[24] NAGARAJ C, HAITCHI H M, HEINEMANN A, et al. Increased Expression of p22phox Mediates Airway Hyperresponsiveness in an Experimental Model of Asthma [J]. Antioxidants & Redox Signaling, 2017, 27(18): 1460-72.DOI:10.1089/ars.2016.6863
[25] SJ BERG L C, NILSSON A Z, LEI Y, et al. Interleukin 33 exacerbates antigen driven airway hyperresponsiveness, inflammation and remodeling in a mouse model of asthma [J]. Scientific Reports, 2017, 7(1): 4219.DOI:10.1038/s41598-017-03674-0
[26] SUNDARAM A, CHEN C, KHALIFEH-SOLTANI A, et al. Targeting integrin α(5)β(1) ameliorates severe airway hyperresponsiveness in experimental asthma [J]. The Journal of Clinical Investigation, 2017, 127(1): 365-74.DOI:10.1172/JCI88555
[27] SAH P K, TEAGUE W G, DEMUTH K A, et al. Poor Asthma Control in Obese Children May Be Overestimated Because of Enhanced Perception of Dyspnea [J]. The journal of allergy and clinical immunology in practice, 2013, 1(1): 39-45.e2.DOI:10.1016/j.jaip.2012.10.006
[28] PERMAUL P, KANCHONGKITTIPHON W, PHIPATANAKUL W. Childhood asthma and obesity—what is the true link? [J]. Annals of allergy, asthma & immunology : official publication of the American College of Allergy, Asthma, & Immunology, 2014, 113(3): 244-6.DOI:10.1016/j.anai.2014.07.001
[29] Passive Smoking Induces Leukotriene Production in Children: Influence of Asthma [J]. Journal of Asthma, 2013, 50(4): 347-53.DOI:10.3109/02770903.2013.773009
[30] COOK J, SAGLANI S. Pathogenesis and prevention strategies of severe asthma exacerbations in children [J]. Current Opinion in Pulmonary Medicine, 2016, 22(1). DOI:10.1097/MCP.0000000000000223
上一篇:
支气管哮喘气道重塑的研究进展
下一篇:
重症哮喘表型研究及治疗策略









